






Welcome to the Affective & Clinical Neuroscience Lab!
We use neuroscientific methods to examine order and disorder in the emotional brain. Specifically, we use neurophysiology and both structural and functional neuroimaging to study the brain systems involved in generating both positive and negative emotions, and the regulation of these systems by the prefrontal cortex.
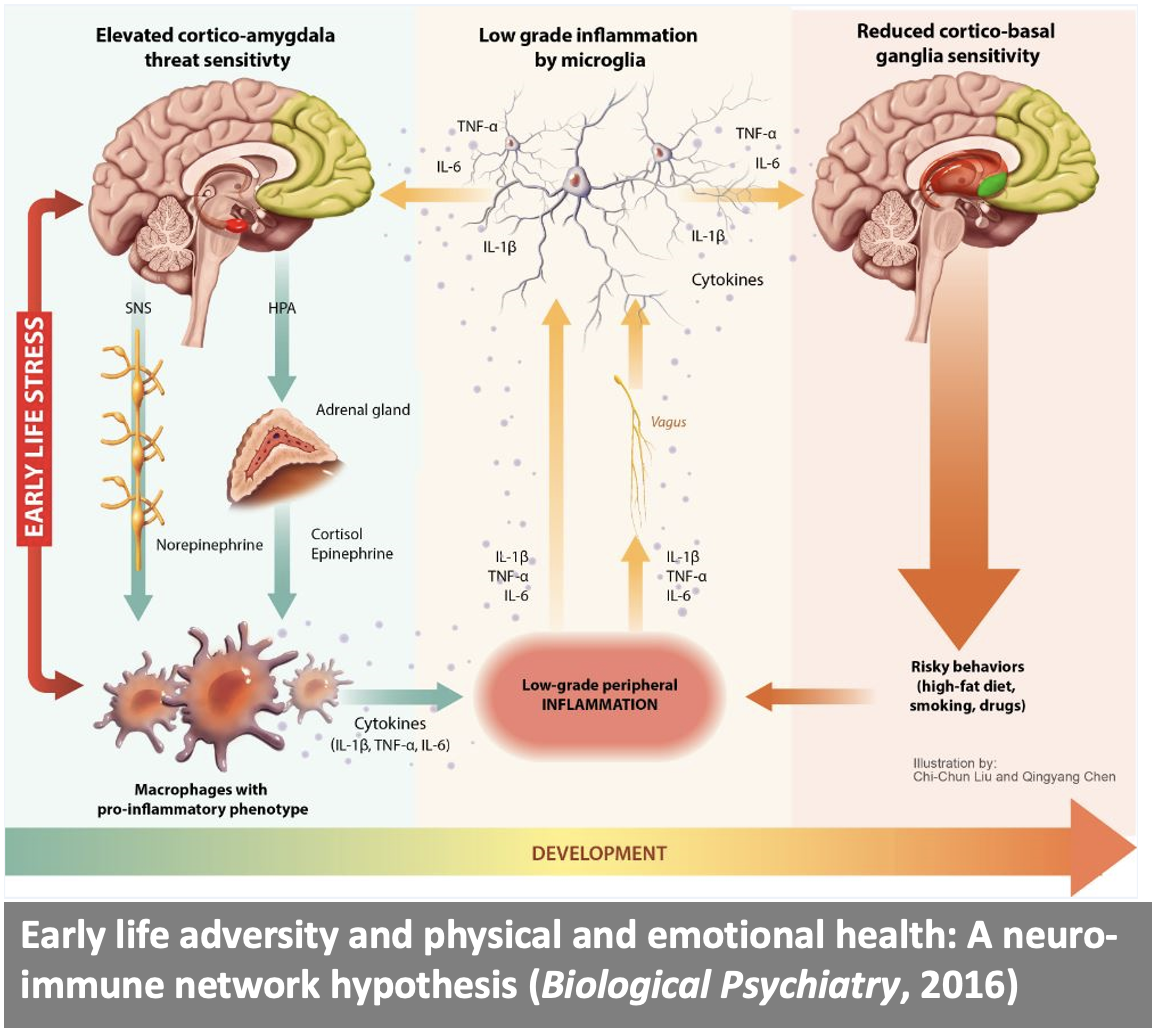
We then aim to translate our research on the emotional brain to investigate the neural mechanisms underlying emotional disorders, including depression, anxiety, addiction, and mania. We also examine how stress affects brain systems involved in generating emotion, and bidirectional signaling between the brain and the immune system in generating risk and resilience for mental and physical health problems.